I. 서론
지난 3개의 글들에서, 구석기 시대 유라시아 인류이동 역사, 유전자학으로 살펴 본 신석기 시대 동북아시아 역사, 그리고 신석기시대 동북아시아의 홍산문화와 앙소문화에 대해 살펴 보았다. (아래 자료 참조). 이번 글은 구석기, 신석기, 청동기, 철기 시대의 동부 유라시아 역사를 전체적으로 종적으로 조명해 보고자 한다. 구석기부터 철기에 이르는 방대한 기간이라서 세밀한 내용을 검토할 수는 없고, 동부 유라시아의 전체적인 흐름을 조명하는 목적으로 글을 쓰고자 한다. 일부 자료는 이미 쓴 상기 내용의 일부를 옯겼다. 본 글은 동북아시아에 12환국이 있었는가에 대한 검토의 일환으로, 자료를 찾아 정리하는 내용이다.
https://paulcgpark.tistory.com/95
구석기시대 유라시아 인류의 이동 역사
I. 서 론 필자는 환단고기가 말하는 12환국이 동북아시아에 없었다는 글을 썼었다. 백두산과의 지형 위치, 그리고 파내류산과 비서갑 등의 위치를 거론하며, 그 가능성이 없다는 것을 주장하였다
paulcgpark.tistory.com
https://paulcgpark.tistory.com/97
유전자학으로 살펴 본 신석기 시대 동북아시아 역사
I. 서론 지난 글에서 구석기시대의 인류이동역사를 살펴 보면서, 12환국시대 이전의 인류 이동역사를 대강 살펴 보았다. (아래 글 참조) https://paulcgpark.tistory.com/95 구석기시대 유라시아 인류의 이
paulcgpark.tistory.com
https://lostcorea.tistory.com/1893
신석기 시대 동북아시아의 홍산문화와 관련된 앙소문화 고찰
I. 서론 지난 두 글에서 구석기 시대의 인류이동 역사와 유전학적으로 살펴 본 신석기시대 동북아시아의 역사를 살펴 보았다. (아래 글 참조). https://lostcorea.tistory.com/1891 12환국 시대 이전 구석기
lostcorea.tistory.com
II. 구석기 시대 동유라시아
1. 구석기 시대의 아무르지역 ANA(Ancient Notheast Asians) 분리
구석기시대의 고대북유라시언(ANE, 남시베리아지역)샘플(24000년전)이나 Tianyuan man 샘플(산동성지역)(4만년전)과 신석시대의 ANA샘플과는 큰 시간적 공백기간이 존재한다. 그러나 아무르지역의 AR19K(19000년전) 샘플은 신석시대의 ANA 유전인자의 토대가 된 것으로 나타난다.
즉 아무르지역의 고대 동북아시아인들(ANA)은 다른 동아시아인들부터 19000년전과 26000년전 사이 어느 시점에서 갈라져 나왔다는 것을 의미한다. (아래 자료 참조)

Amur river in Northeast Asia
"The Prehistoric populations of Eastern Siberia are poorly understood, mainly due to the lack of archaeological specimens. So far, the oldest populations for which genomic data have been obtained are the Upper Paleolithic Ancient North Eurasians (c. 24,000 BP) from Central Siberia, and Upper-Paleolithic populations related to the "Basal-East Asian" Tianyuan man (c. 40,000 BP), specifically the Salkhit (c. 34,000 BP) and AR33K (c. 33,000 BP) samples from Mongolia and the Amur region, or Manchuria. There is then a large gap until the Neolithic period, where the specific ANA gene pool has been identified. Ancestry basal to the ANA gene pool, but significantly closer to them than to the Upper-Paleolithic Tianyuan-related gene pool or other East Asian lineages (such as Southern East Asians), has been found among a sample in the Amur region (AR19K; c. 19 000 BP), suggesting that Ancient Northeast Asians diverged from other East Asian populations sometimes between 19kya to 26kya.[8][9][10] "
(번역) 동시베리아의 선사시대의 인류는 고고학적 자료의 부족으로 매우 빈역하게 이해되고 있다. 현재까지, 유전적 자료로 제일 오래된 인류는 중앙시베리아의 초기 구석기시대의 고대북유라시언(ANE) (24000년전)과 Basal East Asian인 Tianyuan man (4만년전)과 관련된 초기 구석기시대 인류, 그리고 특히, 몽고와 아무르지역 또는 만주지역의 Salkhit표본(34000년전)과 AR33K(33000년전)표본이다. 특별한 ANA 유전자풀이 확인되는 신석기시대까지 큰 시간적 격차가 있다. 초기 구석기시대 Tianyuan man관련 유전자 풀 또는 다른 동아시아계열 (남동아시아인과 같은) 보다 상당히 더 가까운, ANA 유전자풀의 토대가 되는 조상인자가, 아무르지역의 한 샘플(AR19K, 19000년전)에서 발견되었다. 이는 고대 동북아시아인들은 다른 동아시아인들부터 19000년전과 26000년전 사이 어느 시점에서 갈라져 나왔다는 것을 의미한다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
2. 구석기 시대의 ANEA (Ancient Northern East Asians)
Ancient Northern East Asian (ANEA)은 바이칼지역부터 황하강, 그리고 현재 중공의 중앙지역인 Qinling-Huaihe Line까지 퍼져있는 고대 동북아시아인들을 대표하는 혈통이다.(아래 지도 참조). 그들은 약 기원전 20000 - 26000년쯤에 고대 남동아시아인 (Ancient Sourhern East Asians (ASEA)로부터 갈라진 것으로 여겨진다.

Location of the Ancient Northern East Asians[1]
"In archaeogenetics, the term Ancient Northern East Asian (ANEA), also known as Northern East Asian (NEA), is used to summarize the related ancestral components that represent the Ancient Northern East Asian peoples, extending from the Baikal region to the Yellow River and the Qinling-Huaihe Line in present-day central China.[1][3] They are inferred to have diverged from Ancient Southern East Asians (ASEA) around 20,000 to 26,000 BCE.[1][4][5][6]
(번역) 고고학 유전적으로, Ancient Northern East Asian (ANEA)란 단어는, 또한 Northern East Asian (NEA)로도 알려졌는데, 바이칼지역부터 황하강, 그리고 현재 중공의 중앙지역인 Qinling-Huaihe Line까지 퍼져있는 고대 북동아시아인들을 대표하는 조상인자들을 요약한다. 그들은 약 기원전 20000 - 26000년쯤에 고대 남동아시아인 (Ancient Sourhern East Asians (ASEA)로부터 갈라진 것으로 여겨진다.
[출처] <펌>Ancient Northern East Asian (ANEA)|작성자 CG Park
3. 구석기 시대의 ASEA (=Ancient Southern East Asian)
고대 동남아시아인(ASEA)은, Fujian의 Qihe 동굴(12-8천년전), Guangxi(9천년전)뿐만 아니라, Taiwan 해협의 Liangdao 섬 (8천년전) 등의 인류로 대표되는 혈통계열을 말한다. (아래 지도 참조). ASEA조상은 현대의 동아시아, Mainland Southeast Asia, 섬들의 동남아시아, 그리고 오세아니아 등의 유전자 형성에 많이 기여했다.

Approximate distribution of ASEA-affiliated ancestries
"In archaeogenetics, Ancient Southern East Asian (ASEA), also known as Southern East Asian (SEA), is an ancestral lineage that is represented by individuals from Qihe Cave in Fujian (c. 12–8 kya) and Liangdao Island in the Taiwan Strait (c. 8 kya) as well as Guangxi (c. 9 kya). Ancient Southern East Asian ancestry significantly contributed to the genetic makeup of modern populations in East Asia, Mainland Southeast Asia, Insular Southeast Asia, and Oceania, and is commonly associated with the Neolithic expansion of early Austronesian and Austroasiatic speakers that occurred more than 4,000 years ago."
(번역) 고고학 유전자학으로, 고대 동남아시아인(ASEA)은, 남동아시아인(Southern East Asian, SEA)으로도 불리는데, Fujian의 Qihe 동굴(12-8천년전), Guangxi(9천년전)뿐만 아니라, Taiwan 해협의 Liangdao 섬 (8천년전) 등의 인류로 대표되는 혈통계열을 말한다. ASEA조상은 현대의 동아시아, Mainland Southeast Asia, 섬들의 동남아시아, 그리고 오세아니아 등의 유전자구성에 많이 기여했다. 그리고 또한 초기 Austranesian, Austroasiatic 말을 하는 사람들의 신석기 팽창(4천년전 이상)과 공히 연관되어 있다.
초기 Holocene시기까지, Fujian의 고대동남아시아인(ASEA)은 유전적으로 고대동북아시아인(ANEA)과 확연히 구분된다. 이들 ASEA는 다른 동아시아의 고대 동유라시언 계열 (Tianyuan, Hoabinhian, Jomon, Guangxi/Longlin 조상들) 대비하여 확연히 구분된 계통군(clade)을 형성했다. ASEA와 ANEA계열의 분리는 적어도 19000년전에 일어났다.
"Origins
Until the early Holocene, Ancient Southern East Asians from Fujian were genetically clearly distinct from Ancient Northern East Asians (ANEA) who were distributed in an area stretching from the Yellow River to the Amur. The exact origins of both lineages is still only partially understood, but together they formed a distinct clade vis-a-vis all other known ancient East Eurasian lineages in eastern Asia, viz. the Tianyuan, Hoabinhian, Jomon, and Guangxi/Longlin ancestries.[1][2] The split between the ASEA and ANEA lineages must have occurred at least 19,000 years ago, as evidenced by an 19ky-old Upper Pleistocene individual from the Amur river with a clear ANEA affinity.[3] "
(번역) 초기 Holocene시기까지, Fujian의 고대동남아시아인(ASEA)은 유전적으로 고대동북아시아인(ANEA)(황하에서 아무르까지 뻗은 지역에 분포됨)과 확연히 구분된다. 양계열의 정확한 origins은 아직 부분적으로 이해되고 있으나, 이들은 모두 다른 알려진 동아시아의 고대 동유라시언 계열 (Tianyuan, Hoabinhian, Jomon, Guangxi/Longlin 조상들) 대비하여 확연히 구분된 계통군(clade)을 형성했다. ASEA와 ANEA계열의 분리는 적어도 19000년전에 일어났으며, 이는 분명한 ANEA 유사성을 가진 아무르강의 19000년된 초기 구석기인(AR19K)에 의해 증거된다.
중기 Holocene 시기에, ANEA 조상인자를 가진 황하의 수수 경작 농부들의 남쪽으로의 이동은 동아시아 해안의 조상인자 클라인(cline)으로 결과되어 오늘에 이르고 있다. 북쪽의 지나 한족은 대부분 적은 양의 ASEA인자와 ANEA 조상인자를 혼합하여 가지고 있다. 남쪽의 한족과 동남아시아의 비한족 그룹 (즉 Kra-Dai어 사용자와 Hmong-Mien 어 사용자)은 아직도 상당히 높은 수준의 ASEA 조상인자를 가지고 있다. (아래 자료 참조)
"In the mid-Holocene, southward migrations of millet farmers from the Yellow River harboring ANEA ancestry (and also to lesser degree a reverse geneflow of ASEA rice farmers from the Yangtse River to the north) resulted in the coastal East Asian ancestry cline that exists to this day. Northern Han Chinese mostly carry ANEA ancestry with a moderate degree of ASEA admixture, whereas southern Han Chinese as well as non-Han ethnic groups of southern East Asia (viz. speakers of Kra-Dai and Hmong-Mien languages) still carry significantly higher levels of ASEA ancestry.[4][5] "
(번역) 중기 Holocene 시기에, ANEA 조상인자를 가진 황하의 수수 경작 농부들의 남쪽으로의 이동( 그리고 보다 적은 정도로, 양자강의 벼농사 농부인 ASEA 유전인자 소유자의 북쪽으로의 역방향 이동)은 동아시아 해안의 조상인자 클라인(cline)으로 결과되어 오늘에 이르고 있다. 북쪽의 지나 한족은 대부분 적은 양의 ASEA인자와 ANEA 조상인자를 혼합하여 가지고 있다. 남쪽의 한족과 동남아시아의 비한족 그룹 (즉 Kra-Dai어 사용자와 Hmong-Mien 어 사용자)은 아직도 상당히 높은 수준의 ASEA 조상인자를 가지고 있다.
[출처] <펌>Ancient Southern East Asian|작성자 CG Park
III. 신석기시기 동유라시아
신석기 시기의 동유라시아에는 3개의 유전인자가 있다. 즉 ANA(Ancient Northeast Asian), ANEA (Ancient Northern East Asians), ASEA (Ancient Southern East Asians)이 그것이다. 차례대로 살펴 보고자 한다.
1. 신석기시기 ANA (Ancient Northeast Asian)
고고학 유전학적으로, Ancient Northeast Asian(ANA)은, 또한 아무르 조상이라 알려졌는데, 극동러시아, 몽고, 바이칼지역에 7천-4천년전에 살던 수렵인계열을 대표하는 조상유전인자에게 주어진 이름이다. 그들은 약 24000년전에 고대동아시아인 (Ancient East Asians)에서 갈라진 것으로 여겨지며, 알타이산맥 동쪽에서 추출된 고대 인류의 표본에 의해 대표된다. 그들은 고대북동아시아인(Ancient Northern East Asians(ANEA)의 하부그룹이다. (아래 자료 참조)
"In archaeogenetics, the term Ancient Northeast Asian (ANA),[2][3] also known as Amur ancestry,[4] is the name given to an ancestral component that represents the lineage of the hunter-gatherer people of the 7th-4th millennia before present, in far-eastern Siberia, Mongolia and the Baikal regions. They are inferred to have diverged from Ancient East Asians about 24kya ago,[5] and are represented by several ancient human specimens found in archaeological excavations east of the Altai Mountains. They are a sub-group of the Ancient Northern East Asians (ANEA).[6] "
(번역) 고고학 유전학적으로, Ancient Northeast Asian(ANA)은, 또한 아무르 조상이라 알려졌는데, 극동러시아, 몽고, 바이칼지역에 7천-4천년전에 살던 수렵인계열을 대표하는 조상유전인자에게 주어진 이름이다. 그들은 약 24000년전에 고대동아시아인 (Ancient East Asians)에서 갈라진 것으로 여겨지며, 알타이산맥 동쪽에서 추출된 고고학 유물에서 발견된 몇몇 고대 인류의 표본에 의해 대표된다. 그들은 고대북동아시아인(Ancient Northern East Asians(ANEA)의 하부그룹이다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
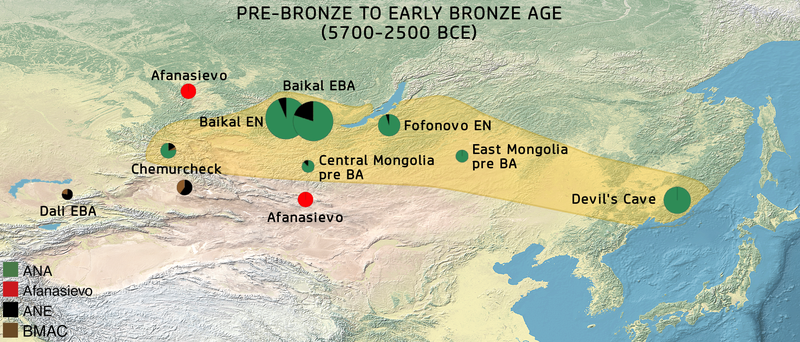
특별한 ANA유전자 그룹으로 인식되는 최초 인류는 극동러시아 태평양가에 있는 Devil's Gate Cave(기원전 5700년)에서 연유한다. ANA/아무르지역 같은 유전자그룹으로서 신석기시대 사람들은 동몽고(BC 4686-4495 BC), 중앙몽고 (BC 3781-3639 BC)에서 확인된다.
Map_of_the_Ancient_Northeast_Asians (ANA) (source : Wikipedia)
Neolithic populations (신석기 인류)
The first individual to be identified with the specific ANA gene pool came from the Russian Far East, near the Pacific coast, at the Devil’s Gate Cave ("DevilsCave_N", c. 5700 BCE).[11] More Neolithic individuals with the ANA/Amur-like gene pool have been identified in eastern Mongolia (SOU001, "EastMongolia_preBA" 4686–4495 cal. BCE), in central Mongolia (ERM003, "CentralMongolia_preBA" 3781–3639 cal. BCE).[12]
(번역) 특별한 ANA유전자 그룹으로 인식되는 최초 개인은 극동러시아 태평양가에 있는 Devil's Gate Cave(기원전 5700년)에서 연유한다. ANA/아무르지역 같은 유전자그룹으로서 신석기시대 사람들은 동몽고(BC 4686-4495 BC), 중앙몽고 (BC 3781-3639 BC)에서 확인된다.
바이칼지역과 시베리아의 인접한 지역의 수렵인 그룹은, 초기 신석기의 동부 바이칼 Fofonovo culture와 서부 바이칼 Kitoi culture (5200-4200 BC)와 연관되어 있다. 뿐만 아니라, 고대 구석기시베리아인(APS)과 함께, Glazkovo culture(2500 BC)와 Cisbaikal_LNBA와 관련있는 초기 청동기시기의 바이칼 인류도 이 수렵인 그룹과 연관되어 있다.
이들 수렵인그룹은 다른 고대북동아시아인 (ANA) 그룹과 광범위하게 무리를 이루었으나, 11% (5-20%)의 작은 고대북유라시아인(ANE) 유전인자 구성에 의해 ANA그룹과 구분된다.
그들은 또한 동북 중공의 Yumin hunter-gatherers (내몽고지방)와 유사한 유전자구성을 보인다. 또한 Yakutia의 신석기와 청동기시기 그룹(Yakutia_LNBA)과 Altai-Sayan 지역의 Krasnoyarsk(kra001)과도 유사한 유전자 구성을 보인다.
이러한 사람들은 때로는 '새로운 시베리아인' (Neo-Siberians)으로 묘사되며, 신석기 Devils Cave표본에 의해 대표되는 ANA/아무르 사람들과도 구분될 수 있다. 그러나 그들의 고대 동북 아시아인 (ANEA)조상을 통해 공통적인 원천을 공유하고 있다. Neo-Siberians은 신석기 아무르조상들의 팽창 이전에 이미 팽창한 것으로 여겨진다.
The closely related hunter-gatherers from the Baikal region and adjacent regions of Siberia are associated with the Early Neolithic eastern Baikal Fofonovo culture ("Fofonovo_EN"), and the western Baikal Kitoi culture ("Baikal_EN", 5200–4200 BCE or Shamanka_EN),[13][14] as well as in conjunction with Ancient Paleo-Siberians (APS), the Early Bronze Age Baikal populations associated with the Glazkovo culture ("Baikal_EBA", circa 2500 BCE or Shamanka_EBA) and Cisbaikal_LNBA.[15][12] They cluster broadly with other Ancient Northeast Asian (ANA) populations, but are differentiated from them via drift associated with an earlier inland expansion route, and a minor Ancient North Eurasians (ANE) component at c. 11% (5-20%).[16][17][18][19] The ANE-like component is best explained via Ancient Paleo-Siberian-rich groups.[20] They also display genetic affinities with the Yumin hunter-gatherers from Northeast China, as well as the Neolithic and Bronze Age groups in Yakutia (Yakutia_LNBA) and Krasnoyarsk (kra001) in the Altai-Sayan region. These populations are sometimes described as "Neo-Siberians" and can be differentiated from proper ANA/Amur populations represented by the Neolithic Devils Cave specimen, but share a common recent origin via their Ancient Northern East Asian ancestor. Neo-Siberians are inferred to have expanded prior to the expansion of Neolithic Amur ancestry.[21][22][23]
(번역) 바이칼지역과 시베리아의 인접한 지역과 긴밀히 연관된 수렵인 그룹은, 초기 신석기의 동부 바이칼 Fofonovo culture와 서부 바이칼 Kitoi culture (5200-4200 BC)와 연관되어 있다. 뿐만 아니라, 고대 구석기시베리아인(APS)과 함께, Glazkovo culture(2500 BC)와 Cisbaikal_LNBA와 관련있는 초기 청동기시기의 바이칼 인류도 이 수렵인 그룹과 연관되어 있다.
이들 수렵인그룹은 다른 고대북동아시아인 (ANA) 그룹과 광범위하게 무리를 이루었으며, 그러나 보다 전의 내륙팽창루트와 관련된 이동과 11% (5-20%)의 작은 고대북유라시아인(ANE) 유전인자 구성에 의해 ANA그룹과 구분된다. ANE와 같은 유전자구성인자는 고대구석기시베리아인(APS) 유전인자가 풍부한 그룹에 의해 잘 설명된다.
그들은 또한 동북 중공의 Yumin hunter-gatherers (내몽고지방)와 유사한 유전자구성을 보인다. 또한 Yakutia의 신석기와 청동기시기 그룹(Yakutia_LNBA)과 Altai-Sayan 지역의 Krasnoyarsk(kra001)과도 유사한 유전자 구성을 보인다. 이러한 사람들은 때로는 '새로운 시베리아인 (Neo-Siberians)으로 묘사되며, 신석기 Devils Cave표본에 의해 대표되는 ANA/아무르 사람들과도 구분될 수 있다. 그러나 그들의 고대 동북 아시아인 (ANEA)조상을 통해 공통적인 원천을 공유하고 있다. Neo-Siberians은 신석기 아무르조상들의 팽창 이전에 이미 팽창한 것으로 여겨진다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
Devils 동굴샘플은 아무르지역의 14000년전의 샘플 (AR14K)과 유전적으로 이어지는 것을 보여 주고 있다. 이는 ANA의 특별한 유전자풀이 14000년전부터 형성되었음을 말해준다. 신석기시대의 ANA유물은 서쪽으로 1500km 떨어진 알타이산맥에서까지 발견되었다.
- Devils Cave sample
The Devils_Cave_N sample was found to display genetic continuity with a c. 14kya old sample (AR14K) from the Amur region, suggesting that the specific ANA gene pool formed as early as 14,000 BP.[9] Neolithic ANA remains have been found as far as the Altai Mountains, 1,500 km further to the west than previously understood.[24]
(번역) Devils 동굴 N 샘플은 아무르지역의 14000년전의 오래된 샘플 (AR14K)과 유전적으로 이어지는 것을 보여 주고 있다. 이는 ANA의 특별한 유전자풀이 14000년전부터 형성되었음을 말해준다. 신석기시대의 ANA유물은 이미 이해된 것보다 더 서쪽으로 1500km 떨어진 알타이산맥에서까지 발견되었다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
2. 신석기 시대의 ANEA (Ancient Northern East Asians)
ANEA (Ancient Northern East Asian)는 3개의 하위그룹으로 나눌 수 있다: 즉 ANA (Ancient Northeast Asians), Neo-Siberians (신 시베리아인), 그리고 황하농민들 (Yellow River farmers) 들이다.
ANA는 다른 말로 '아무르조상'으로 알려져 있는데, ANEA그룹의 한 하위그룹을 이루며, 특히 7000-4000년전의 수렵인그룹의 조상이다. 이들은 활동지역이 아무르지역 그리고 후에 동시베리아, 몽골, 바이칼지역에 더 퍼졌으며, 다른 고대 북쪽의 동아시아인, 예를 들어 초기 신석기 바이칼지역에 퍼진 신시베리아인 등에 가장 가깝게 연계되어 있다.
"The ANEA can be differentiated into broadly three sub-groups, namely the “Ancient Northeast Asians“ (ANA), “Neo-Siberians", and "Yellow River farmers". The ANEA are to be distinguished from the namely similar "Ancient Northeast Asian" (ANA) lineage, which is alternatively also known as "Amur ancestry", and which forms a sub-group of the ANEA grouping, specifically ancestral to hunter-gatherer people of the 7th-4th millennia before present, in the Amur region and later expanding to far-eastern Siberia, Mongolia and the Baikal regions, but which are most closely related to other ancient northern East Asians, such as the earlier expanding "Neo-Sibersians" evident in the Early Neolithic Baikal region.[2][7]
(번역) ANEA (Ancient Northern East Asian)는, 차이가 있지만, 대강 3개의 하위그룹으로 나눌 수 있다: 즉 ANA (Ancient Northeast Asians), Neo-Siberians (신 시베리아인), 그리고 황하농민들 (Yellow River farmers) 들이다. ANEA는 유사한 ANA계열과 구분되는데, ANA는 다른 말로 '아무르조상'으로 알려져 있는데, ANEA그룹의 한 하위그룹을 이루며, 특히 7000-4000년전의 수렵인그룹의 조상이다. 이들은 활동지역이 아무르지역 그리고 후에 동시베리아, 몽골, 바이칼지역에 더 퍼졌으며, 다른 고대 북동아시아인 (예를 들어 초기 신석기 바이칼지역에 퍼진 신시베리아인) 등에 가장 가깝게 연계되어 있다.
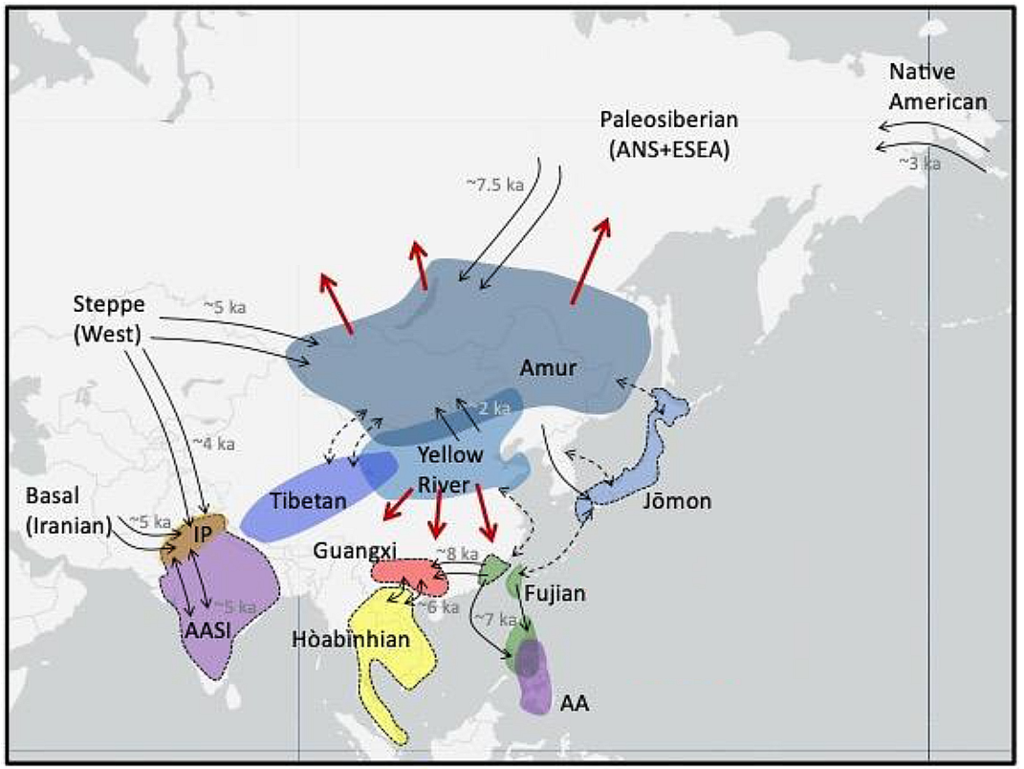
Location of the major sub-groups within the Ancient Northern East Asians, with the Yellow River farmers in the Yellow River valley, and Ancient Northeast Asians (or Amur ancestry) above[2]
ANEA계열의 초기 집단은, 북으로 고대북유라시언(ANE)과 함께 고대구석기시베리아인(APS)을 형성하는데 이바지한 것으로 여겨진다. 남으로는 ASEA(고대남동아시아인)과 함께 "황하의 농민들"(Yellow River farmers)을 형성하는데 이바지 했다.
"To the north, an early branch of the Ancient Northern East Asian lineage is inferred to have contributed to the formation of the Ancient Paleo-Siberians (APS) in conjunction with the Ancient North Eurasians (ANE), and, in the south, to the formation of the "Yellow River farmers" in conjunction with the Ancient Southern East Asians (ASEA).[1][4] Yellow River farmers are associated with the spread of Sino-Tibetan languages.[8]
(번역) 북으로는, ANEA계열의 초기 집단이, 고대북유라시언(ANE)과 함께 고대구석기시베리아인(APS)을 형성하는데 이바지한 것으로 여겨진다. 남으로는 ASEA(고대남동아시아인)과 함께 "황하의 농민들"(Yellow River farmers)을 형성하는데 이바지 했다. 황하의 농민그룹은 Sino-Tibetan 언어들의 전파와 연관되어 있다.
"Neo-Siberians"은, ANEA(14000년전)의 내륙확장루트와 연관 있으며, 그러나 "ANA조상"과 관련 있는 Amur-hunter-gatherers(7000-14000년전)와는 다를 수 있다.
이 Neo-Siberians은 우선적으로 신석기, 청동기시기의 바이칼지역그룹의 조상이 되었다.
이들은 초기의 범 우랄어사용자들의 팽창과 연관 있을 수 있으며, APS와 더불어, 청동기 시기 서부 바이칼 수렵인 그룹 (초기 예니세이 언어 사용자와 관련 있는 Glazkovo culture(2500 BC)과 Cisbaikal_LNBA조상과 관련 있음)과도 연관 있을 수 있다.
"The "Neo-Siberians" or "inland Northeast Asians", represented by the Yumin hunter-gatherers and Transbaikal_EMN ancestry, are associated with an inland expansion route of Ancient Northern East Asians (China_NEastAsia_Inland_EN, c. 14kya), but which can be differentiated from the "Amur hunter-gatherers" (c. 7-14kya) associated with "ANA ancestry". This branch became primarily ancestral to Neolithic and Bronze Age groups in the Baikal region, such as the Neolithic Baikal hunter-gatherers from the Kitoi culture ("Baikal_EN", 5200–4200 BCE or Shamanka_EN), the Late Neolithic/Bronze Age Yakutia (Yakutia_LNBA) and Krasnoyarsk (kra001_BA) ancestry in Eastern Siberia and the Altai-Sayan region, which may be associated with the expansion of early Proto-Uralic speakers, as well as, in conjunction with Ancient Paleo-Siberians, to the Bronze Age Western Baikal hunter-gatherers associated with the Glazkovo culture ("Baikal_EBA", circa 2500 BCE or Shamanka_EBA) and Cisbaikal_LNBA ancestry, which may be associated with early Yeniseian speakers.[9][10][11][12]
(번역) Yumin hunter-gatheres와 Transbaikal_EMN 조상에 의해 대표되는 "Neo-Siberians" 또는 내륙의 동북아시아인 ("Inland Northeast Asians")은, ANEA(14000년전)의 내륙확장루트와 연관 있으며, 그러나 "ANA조상"과 관련 있는 Amur-hunter-gatherers(7000-14000년전)와는 다를 수 있다. 이 Neo-Siberians은 우선적으로 신석기 청동기시기의 바이칼지역그룹 - Kitoi culture(5200-4200 BC)의 신석기 바이칼 수렵인그룹과, 동시베리아와 알타이 사이언지역에서의 말기 신석기/청동기 시기 Yakutia와 krasnoyarsk 조상들 - 의 조상이 되었다. 이들은 초기의 범 우랄어사용자들의 팽창과 연관 있을 수 있으며, APS와 더불어, 청동기 시기 서부 바이칼 수렵인 그룹 (초기 예니세이 언어 사용자와 관련 있는 Glazkovo culture(2500 BC)과 Cisbaikal_LNBA조상과 관련 있음)과도 연관 있을 수 있다.
고대 동북아시아인(ANA)(아무르조상)은 "Neo-Siberians"그룹이 흩어진 후 팽창했으며, 튀르크, 몽골, 퉁구스어 사용자들의 확산과 관련되어 있을 수 있다.
"Acient Northeast Asians (Amur ancestry), represented by Mesolithic Amur specimens (c. 7-14kya) and subsequent samples from Mongolia, expanded after the dispersal of "Neo-Siberian" like groups, and may be associated with the spread of Turkic, Mongolic, and Tungusic speakers.[9][11]
(번역) 고대 동북아시아인(ANS)(아무르조상)은 중석기시기 아무르 표본(7000-14000년전)과 몽고의 뒤이은 샘플에 의해 대표되는데, 이들은 "Neo-Siberians" 같은 그룹이 흩어진 후 팽창했으며, 튀르크, 몽골, 퉁구스어 사용자들의 확산과 관련되어 있을지도 모른다.
Paleolithic and Neolithic specimens
The ANEA lineage is represented by a late Paleolithic specimen (c. 19kya) from the Amur region (Amur19k), as well as Early Neolithic samples including the Yumin, Devil's Gate (Far East Russia, ~7.7 kya), Shandong (coastal China, ~9.5-7.5 kya) and Lake Baikal (southern Siberia, ~7.1-6.3 kya) individuals.[1][13]
(번역) 구석기 및 신석기 표본들
ANEA계열은 아무르지역의 후기구석기시대 표본 (AR19K)에 의해 대표된다. 또한 초기 신석기시대 샘플들, 예를 들어, Yumin, Devil's Gate (극동러시아, 7700년전), 산동지역(중공 해안가, 9500-7500년전), 그리고 바이칼호수(남시베리아, 7100-6300년전)의 표본들도 ANEA계열을 대표한다.
[출처] <펌>Ancient Northern East Asian (ANEA)|작성자 CG Park
3. 신석기 시대의 ASEA (=Ancient Southern East Asian)
Neolithic expansion into Southeast Asia and Oceania
(동남아시아와 오세아니아로의 신석기시대 팽창)
기원전 3천년대부터, 쌀농사 기반의 농업은 남동아시아에서 중국 대륙 및 섬지역의 동남아시아로 퍼져 나갔다. 이러한 기술적 전파는 ASEA 조상인자를 가진 남동아시아 농업인들의 이주의 결과이다. 이러한 신석기 농부들은 두가지 길로 이동했다: 중국 본토 남동아시아로 향하는 내륙의 길, 그리고 타이완을 발생지로 하는 해상루트이다.
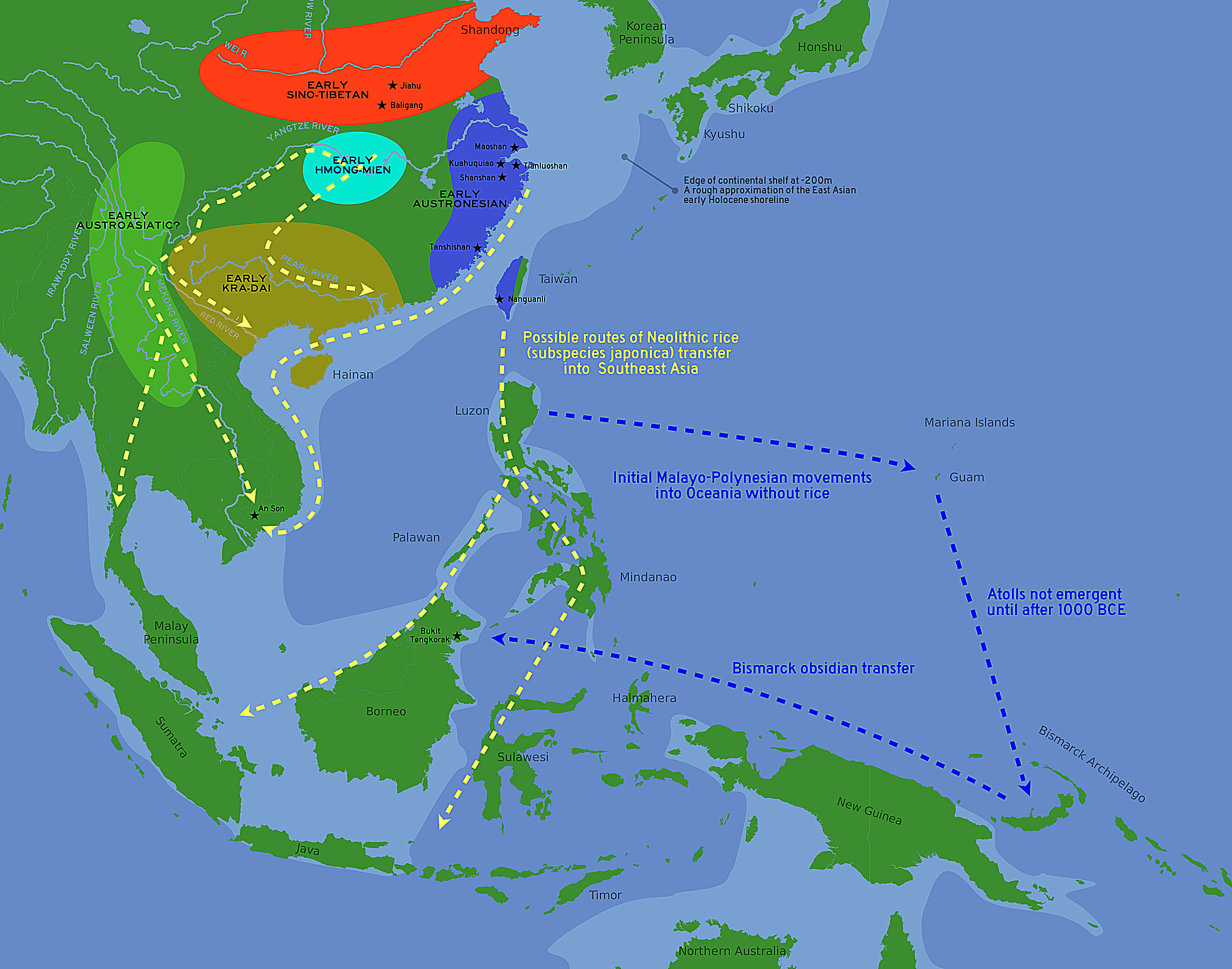
Possible language family homelands and routes of early rice transfer.
"Starting from the third millennium BCE, rice farming-based agriculture spread from southern East Asia into Mainland and Insular Southeast Asia. This technological spread was a result of the migration of southern East Asian agriculturalists that carried ASEA ancestry. These Neolithic farmers took two routes: an inland route into Mainland Southeast Asia, and an maritime route that originated from Taiwan.[6][5][7]
(번역) 기원전 3천년대부터 시작하여, 쌀농사 기반의 농업은 남동아시아에서 중국 대륙 및 섬지역의 동남아시아로 퍼져 나갔다. 이러한 기술적 전파는 ASEA 조상인자를 가진 남동아시아 농업인들의 이주의 결과이다. 이러한 신석기 농부들은 두가지 길로 이동했다: 중국 본토 남동아시아로 향하는 내륙의 길, 그리고 타이완을 발생지로 하는 해상루트이다.
4천년전의 대륙 남동아시아의 최초 농부들의 고대 DNA는 ASEA계열의 혈통으로부터 비롯되는데, 지역적인 수렵인들과 상당한 혼합의 모습이다. 이 신석기 대륙의 남동아시아 조상인자는 Austroasiatic어를 사용하는 동남아시아의 그룹(특히 라오스와 타일란드의 Mlabri와 Htin 지역인들)과 동아시아, 남아시아 일부의 현대인들에게서 절정을 이룬다. 여기서, 동남아대륙의 농업의 전파는 Austroasiatic언어 사용자들의 팽창과 광범위하게 연계되어 있다.
"Ancient DNA of first farmer individuals from Mainland Southeast Asia dated at c. 4kya derives most of its ancestry from the ASEA lineage, with significant admixture from a local hunter-gatherer population.[a] This Neolithic Mainland Southeast Asian ancestry peaks among modern populations in Austroasiatic-speaking groups of Southeast Asia (most notably in the Mlabri and Htin peoples in northern Laos and Thailand) and parts of East Asia and South Asia. Hence, the first spread of farming in Mainland Southeast Asia is widely assumed to be linked to the expansion of the Austroasiatic languages.[8][6] From Mainland Southeast Asia, this Austroasiatic-related ancestry spread into Insular Southeast Asia to the Sunda Islands,[8] adjacent areas (viz. Palawan, Mindanao) of the Philippines,[11] and western Wallacea,[12][13] although there are no remaining Austroasiatic languages spoken in this area, having been supplanted by incoming Austronesian languages.
(번역) 4천년전의 본토 남동아시아의 최초 농부들의 고대 DNA는 ASEA계열의 조상인자로부터 비롯되는데, 지역적인 수렵인들과 상당한 혼합의 모습이다. 이 신석기 대륙의 남동아시아 조상인자는 Austroasiatic어를 사용하는 동남아시아의 그룹(특히 라오스와 타일란드의 Mlabri와 Htin 지역인들)과 동아시아, 남아시아 일부의 현대인들에게서 절정을 이룬다. 여기서, 동남아대륙의 농업의 최초 전파는 Austroasiatic언어 사용자들의 팽창과 광범위하게 연계되어 있다. 동남아 대륙으로부터, 이 Austroasiatic관련 조상인자는 섬들의 동남아시아와 순다랜드, 필리핀의 인접지역(즉 Palawan, Mindanao), 서부 Wallacea로 퍼졌다. 그러나 지금은 이 지역에 Austroasiatic언어는 남아 있지 않고, Austronesian 언어로 대체되었다.
5천-4천년전부터 시작한 초기 Austronesians의 해상을 통한 팽창은 타이완부터 필리핀, 인도네시아 군도, 오세아니아로 ASEA 조상인자가 퍼졌다. 이후 Austronesian 언어 사용자들의 이동은 ASEA조상인자를 멀리 마다가스카르와 동부 폴리네시아까지 이동시켰다.
"The rapid maritime expansion of the early Austronesians starting c. 5,000–4,000 years ago brought ASEA ancestry from Taiwan to the Philippines, the Indonesian archipelago and Oceania, initially with little admixture from local populations, as can be seen from 2,900–2,500 year-old Lapita-related individuals from Vanuatu and Tonga,[14] and from ancient 2,800–2,200 year-old DNA of the first settlers of Guam.[15] In western Indonesia, Austronesian settlers admixed with people from the Austroasiatic-related settlement stream with Neolithic Mainland Southeast Asian ancestry,[5] while in eastern Indonesia and Oceania, all Austronesian-speaking groups have Papuan-related geneflow at various levels.[14][12][15] Later migrations of Austronesian speakers brought ASEA ancestry as far as to Madagascar and eastern Polynesia.[6]
(번역) 5천-4천년전부터 시작한 초기 Austronesians의 해상을 통한 팽창은 타이완부터 필리핀, 인도네시아 군도, 오세아니아로 ASEA 조상인자가 퍼졌다. 그러나 지역 주민들과는 거의 혼합되지 않았다. 이는 Vanuatu와 Tonga의 2900-2500년된 Lapita-관련된 인류에서 볼 수 있으며, 괌의 최초 정착인의 2800-2200년된 DNA에서 볼 수 있다. 서부 인도네시아의 Austronesians정착인들은 신석기 대륙 동남아시아 조상인자를 가진 Austroasiatic 관계된 정착흐름과 관련된 사람들과 섞였다. 반면, 동부 인도네시아와 오세아니아에서는 모든 Austronesian어 사용자그룹은 여러 수준에서 Papuan관련 유전자를 가지고 있다. 이후 Austronesian 언어 사용자들의 이동은 ASEA조상인자를 멀리 마다가스카르와 동부 폴리네시아까지 이동시켰다.
[출처] <펌>Ancient Southern East Asian|작성자 CG Park
IV. 청동기 시기 동유라시아
1. Ulaanzuukh and Slab Grave cultures
Ulaanzuukh(1450-1150 BC)와 Slab Grave (1100-300 BC) 문화의 사람들은 ANA(아무르 조상인자)의 직접 자손들로 정형화할 수 있다. 그들은 이전의 신석기 및 초기 청동기시기의 바이칼 수렵인들을 광범위하게 대체했다. 그들 사이의 유전적 교류가 있었는데, 특히, 원래 Amur-N과 같은 혈통을 가진 신석기 동부 몽골인들과 지역의 바이칼 수렵인들 사이의 유전적 교류가 있었다 한다.
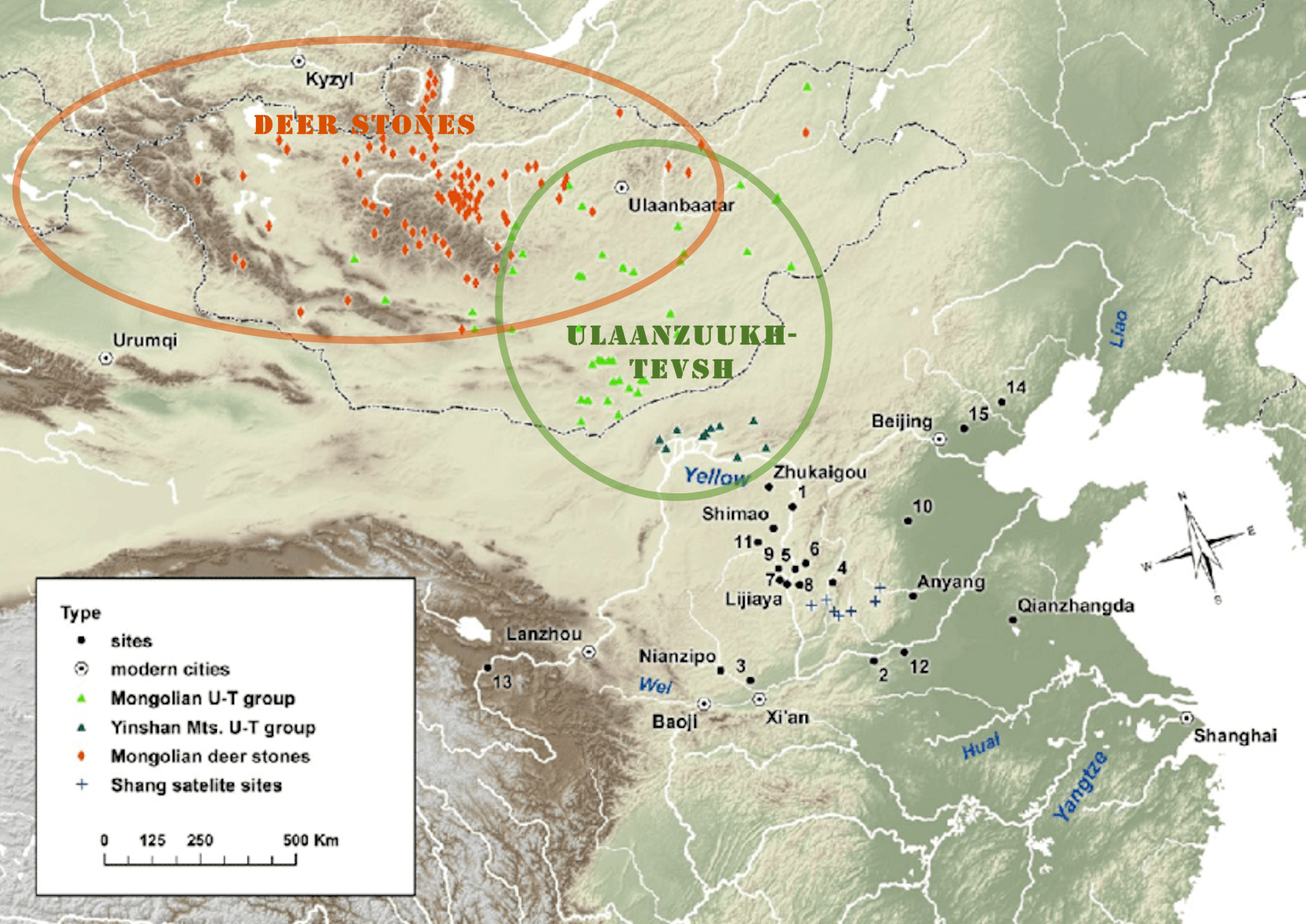
Ulaannzuukh - Tevsh sites (source : Wikipedia)

Slab Grave culture area (source : Wikipedia)
"The people of the Ulaanzuukh (1450–1150 BCE) and Slab Grave (1100–300 BCE) cultures were closely associated with the Ancient Northeast Asians (Amur ancestry) and can be modeled as direct descendants of them. They largely replaced the previous Neolithic and Early Bronze Age Baikal hunter-gatherers, although geneflow between them has been proposed, particularly between a Neolithic Eastern Mongolian population (East_Mongolia_preBA) having primarily Amur_N-like ancestry and local Baikal hunter-gatherers (Baikal_EBA).[25]
(번역) Ulaanzuukh(1450-1150 BC)와 Slab Grave (1100-300 BC) 문화의 사람들은 ANA(아무르 조상인자)와 매우 밀접하게 연관되어 있으며, 이들의 직접 자손들로 정형화할 수 있다. 그들은 이전의 신석기 및 초기 청동기시기의 바이칼 수렵인들을 광흐범위하게 대체했다. 비록 그들 사이의 유전적 교류가 제시되었지만 말이다. 특히, 원래 Amur-N과 같은 조상인자를 가진 신석기 동부 몽골인들 (East_Mongolia_preBA)과 지역의 바이칼 수렵인들(Baikal_EBA) 사이의 유전적 교류가 있었다 한다.
신석기 및 초기 청동기시기 바이칼 수렵인 후계 그룹은 서부초원유목민(Western Stepple Herders)/Sintashta 문화 같은 이들과 다양한 정도의 혼합이 후기 청동기시기동안에 알타이지역에서 나타나기 시작했다. 이러한 그룹은 동부초원으로부터 신석기 및 초기 청동기 바이칼지역인들로부터 형성되었다. 그리고 서부초원유목민(WSH) 이주그룹과의 연이은 혼합이 이루어졌다. 이것은 북몽골의 Khovsgol LBA (Late Bronze Age)유목민과 알타이지역의 Altai MLBA (Middle Late Bronze Age)수렵인그룹을 포함한다.
"Several successor groups of the Neolithic and Early Bronze Age Baikal hunter-gatherers with varying degrees of Western Steppe Herders/Sintashta-like admixture started to appear in the Altai region during the Late Bronze Age. These groups formed from the Neolithic and Early Bronze Age Baikal populations from the Eastern Steppe and subsequent admixture from Western Steppe Herder migrant groups. This includes the Khövsgöl LBA[26] herders from northern Mongolia and the Altai MLBA hunter-gatherers from the Altai region.[27][12]
(번역) 신석기 및 초기 청동기 시기 바이칼 수렵인의 몇몇 후계 그룹은 서부초원유목민(Western Stepple Herders)/Sintashta 문화 같은 이들과 다양한 정도의 혼합이 후기 청동기시기동안에 알타이지역에서 나타나기 시작했다. 이러한 그룹은 동부초원으로부터 신석기 및 초기 청동기 바이칼지역인들로부터 형성되었다. 그리고 서부초원유목민(WSH) 이주그룹과의 연이은 혼합이 이루어졌다. 이것은 북몽골의 Khovsgol LBA (Late Bronze Age)유목민과 알타이지역의 Altai MLBA (Middle Late Bronze Age)수렵인그룹을 포함한다.
Khovsgol 후기청동기 유목민들은 대부분의 초기 청동기 바이칼 수렵인그룹(Baikal EBA or Shamanka EBA, 93-96%)과 약간의 서부초원유목민(WSH) (Sintashta, 4-7%)들의 혼합의 자손들이다. 유전적 분석은, 이들이 서부초원유목민들(WSH)로부터 낙농유목이 채택되었고, 그들은 원래 지역의 동북아시아 출신임이 밝혀졌는데, 이는 문명적 전파를 의미한다
"The Khövsgöl LBA herders are descended from Early Bronze Age Baikal hunter-gatherers (Baikal EBA or Shamanka EBA, c. 93-96%) with small amounts of admixture from Western Steppe Herders (Sintashta, c. 4-7%). Genetic analyses revealed that while dairy pastoralism seems to have been adopted by them from the Western Steppe Herders, they were primarily of local Northern East Asian origin, implying cultural transmission. Modern day Tuvans and Nganasans, followed by Nanais, Yukaghirs, Evens, Itelmens, Ulchis, Koryaks, Nivkhs, and Chukchis, are among the people sharing the highest genetic affinities with the Late Bronze Age herders of Khövsgöl, but are not identical with them.[28][29][30][19]
(번역) Khovsgol LBA(후기청동기) 유목민들은 대부분의 초기 청동기 바이칼 수렵인그룹(Baikal EBA or Shamanka EBA, 93-96%)과 약간의 서부초원유목민(WSH) (Sintashta, 4-7%) 혼합의 자손들이다. 유전적 분석은, 이들이 서부초원유목민들(WSH)로부터 낙농유목이 채택되었고, 그들은 원래 지역의 동북아시아 오리진임이 밝혀졌는데, 이는 문명적 전파를 의미한다. 현대의 Tuvans , Nganasans, Nanais, Yukaghirs, Evens, Itelmens, Ulchis, Koryaks, Nivkhs, and Chukchis 들은 후기청동기시기의 Khovsgol유목민과 매우 높은 유전적 유사성을 공유한 사람들사이에 있으며, 그러나 동일한 것은 아니다.
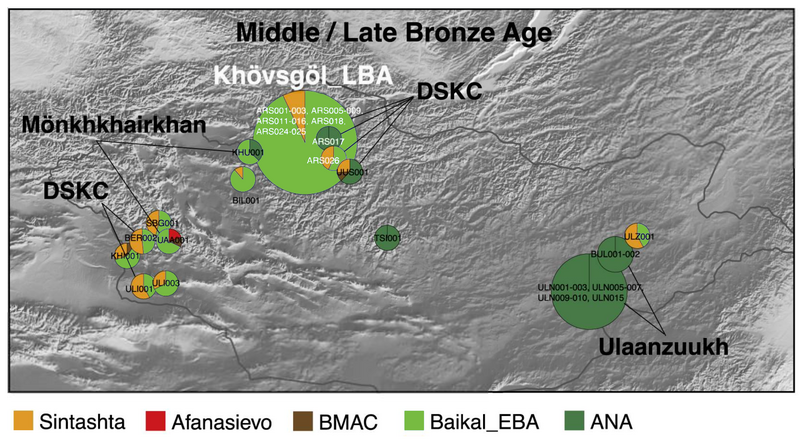
Middle-Late Bronze Age genetics in Mongolia
더 서쪽의 알타이 중기후기청동기시기 유전적 그룹은, 동부 스키타인(사카) (바이칼 /Shamanka EBA그룹의 40-55% 조상인자를 가진 그룹)과 연관되어 있으며, 또한 나머지부분은 초기 인도이란인과 관련있는 Sintashta같은 그룹과 혼합 (45-60%)되어 있다.
"The Altai MLBA gene pool further West can be associated with Eastern Scythians (Saka), who can be modeled as deriving significant amounts of ancestry (c. 40-55%) from the Baikal/Shamanka EBA groups, with the remainder being derived from Sintashta-like admixture (c. 45-60%) associated with early Indo-Iranians.[31][32]
(번역) 더 서쪽의 알타이 중기후기청동기시기 유전적 그룹은, 동부 스키타인(사카) - 바이칼/Shamanka EBA그룹의 40-55% 조상인자를 가진 그룹 - 과 연관되어 있으며, 또한 나머지부분은 초기 인도이란인과 관련있는 Sintashta같은 그룹과 혼합 (45-60%)에 관련되어 있다.
타림 미이라(기원전 2000년)는 높은 수준의 ANE(고대 북유라시언) 조상인자(72%)와 보다 적은 동아시아인구 (특히 Baikal_EBA 28%)의 혼합을 보여 주었다.
"Tarim Mummies (타림 미이라)
A genomic study published in 2021 found that the Tarim mummies (c. 2000 BCE) had high levels of Ancient North Eurasian ancestry (c. 72%), with a smaller admixture from an East Asian-like population (particularly the Baikal_EBA, at c. 28%), but no detectable Western Steppe Herder-related ancestry.[35][36]
(번역) 2021년 발표된 유전자 연구에 의하면, 타림 미이라(기원전 2000년)는 높은 수준의 ANE(고대 북유라시언) 조상인자(72%)와 보다 적은 동아시아인구 (특히 Baikal_EBA 28%)의 혼합을 보여 주었다. 그러나 Western Steppe Herder(서부초원수렵인) 관련 조상인자는 발견되지 않았다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
V. 철기 시대 동유라시아
1. Sakas, Xiongnus, Huns, Avars (사카, 흉노, 훈족, 아바르)
바이칼 초기 청동기 인류는 혼종의 유라시언 Scytho-Siberians cultures의 형성에 지대한 공헌(40-55%)을 하였다. 이들 Scytho-Siberians cultures는 BC 1000년경부터 중앙아시아의 Tasmola문화 (Central Saka) 뿐만 아니라, Arzhan, Pazyryk culture (동부 사카)들을 말한다. (아래 지도 참조)
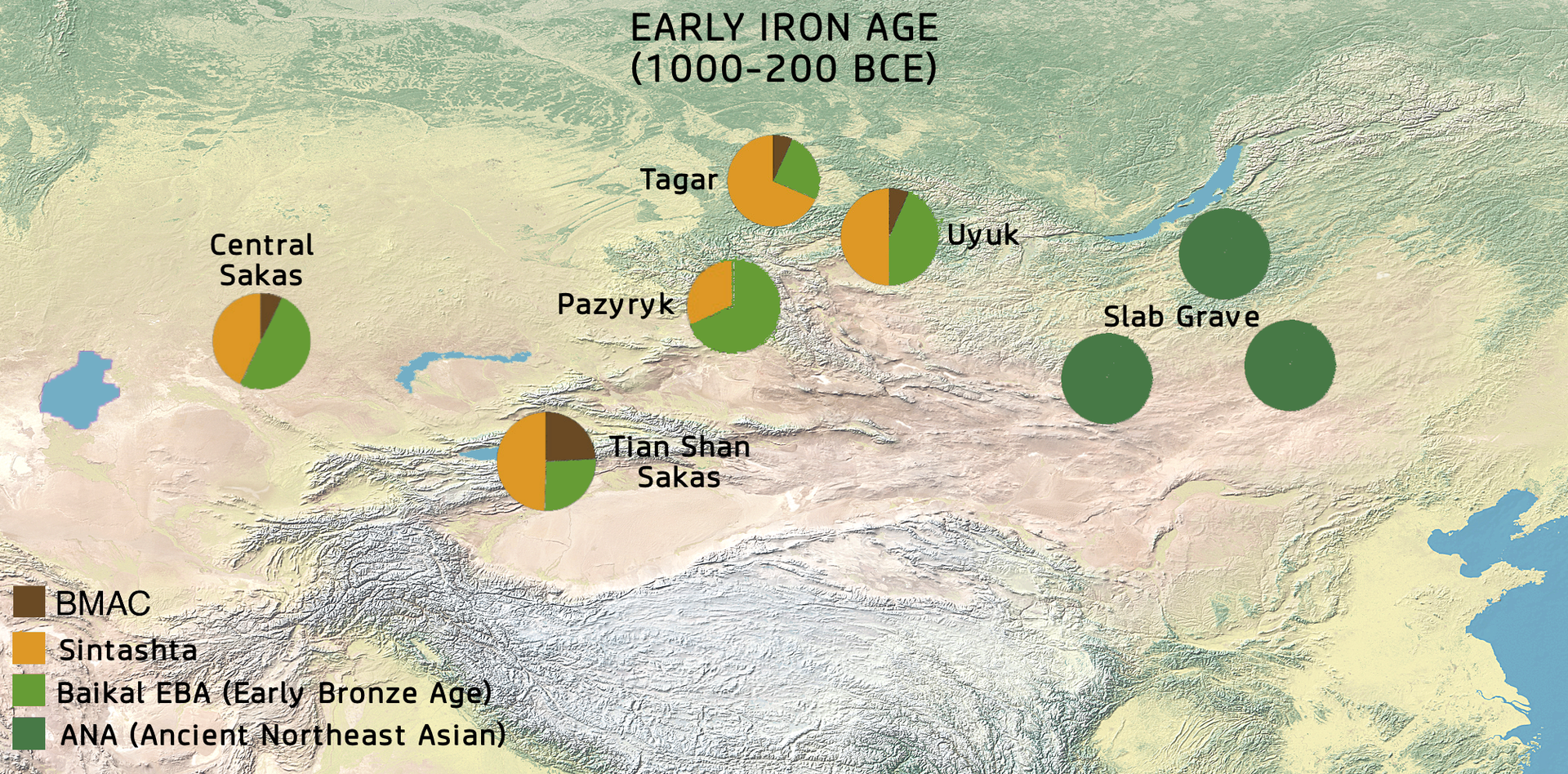
The Slab Grave culture was essentially Ancient Northeast Asian (ANA, ), while the neighbouring Sakas combined in almost equal parts Western Eurasian (Sintashta, ) with Ancient Northeast Asian (Baikal EBA, ) ancestry, with a smaller Iranian contribution (BMAC, ).[37]
"he Baikal EBA populations, also contributed to a large extent to the formation of the hybrid Eurasian Scytho-Siberian cultures, such as the Arzhan and Pazyryk (Eastern Saka) as well as the Tasmola (Central Saka) cultures of Central Asia from around 1,000 BCE, contributing about half of their genetic profile (40-55%), highlighting the increase in genetic diversity during the late Bronze Age and the following Iron Age.[37]
(번역) 바이칼 초기청동기 인류는 혼종의 유라시언 Scytho-Siberians cultures - BC 1000년경부터 중앙아시아의 Tasmola문화 (Central Saka) 뿐만 아니라, Arzhan, Pazyryk culture (동부 사카) - 의 형성에 지대한 공헌을 하였다. 그들의 유전적 프로필의 반이상 (40-55%)을 형성하는데 이바지했으며, 이는 후기 청동기와 뒤이은 철기시대에 유전적 다양성이 증가하는 것을 강조한다.
혼종의 사카문화는 흉노제국 (기원전 3세기-기원후 1세기)의 형성에 중요한 역할을 했다. 이 흉노제국은 특별한 사카 혈통(특히 Chandman/Uyuk같은 것들)과 신석기 아무르 혈통 (Ulaanzuukh 와 Slab Grave 혈통들)을 합한 것이다. (필자 주 : 여기서 흉노제국은 중공사학자들이 주장하는 중공 북방의 민족을 말한다.) 이후에는 Sarmatian(동유럽)과 한족 혈통이 더 추가되었다.
흉노(=중공 북방유목민)의 상위 계층은 유전적 동질성이 보다 많으며, 그들의 혈통은 주로 동유라시언 (Ulaanzuukh/Slab Grave culture)으로부터 나왔다. 반면 하위계층의 사람들은 보다 다양하고 높은 바율의 사카 혈통을 가지는 경향이 있다.
제국의 남자리더, 'Chanyu (선우)' 같은 사람은 서쪽 국경의 고위계층 여인과 유사한 혈통을 가진 것으로 나타난다. (중공북방의 흉노 유목민은) 39.3%는 Slab Grave 유전 혈통, 51.9%의 Han (중공지역 한족)혈통, 그리고 나머지 8.8%의 사카 (Chandmann) 혈통을 나타낸다.
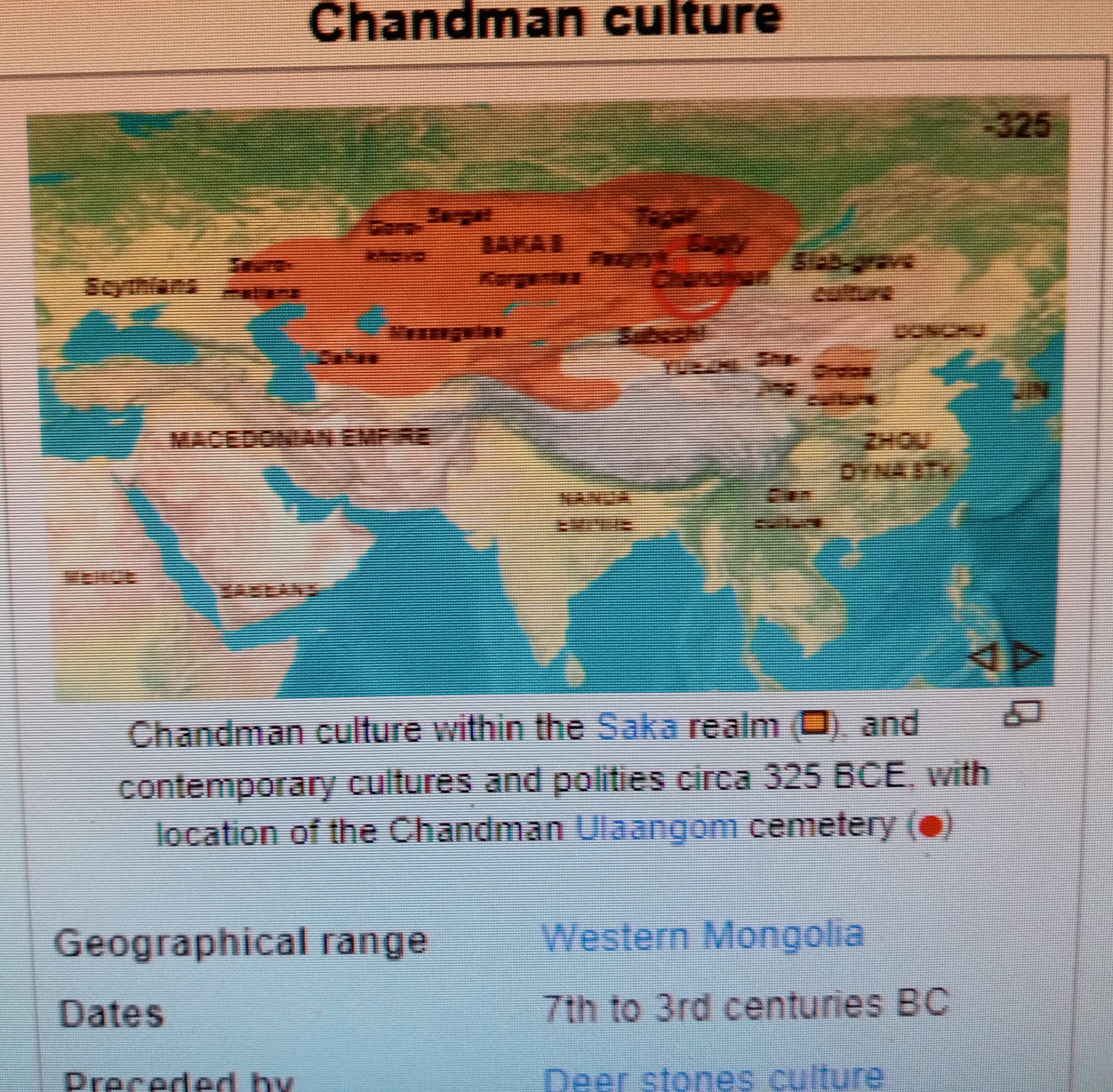
Chandman culture location in Saka realm (source : Wikipedia)
The hybrid Saka cultures in turn played an important role in the formation of the Xiongnu Empire (3rd century BCE-1st century CE), which combined specific Saka ancestries (particularly Chandman/Uyuk-related ones), with Neolithic Amur-derived Ulaanzuukh and Slab Grave ancestries, to which Sarmatian and Han ancestry was further added at a later stage.[37] High status Xiongnu individuals tended to have less genetic diversity, and their ancestry was essentially derived from the Eastern Eurasian Ulaanzuukh/Slab Grave culture, while low status individuals tended to be more diverse and having higher Saka-like ancestry.[39] A likely chanyu, a male ruler of the Empire identified by his prestigious tomb, was shown to have had similar ancestry as a high status female in the "western frontiers", deriving about 39.3% Slab Grave genetic ancestry, 51.9% Han ancestry, with the rest (8.8%) being Saka (Chandman) ancestry.[39]
(번역) 혼종의 사카문화는 역으로 흉노제국 (기원전 3세기-기원후 1세기)의 형성에 중요한 역할을 했다. 이 흉노제국은 특별한 사카 혈통(특히 Chandman/Uyuk같은 것들)과 신석기 아무르 혈통 (Ulaanzuukh 와 Slab Grave 혈통들)을 합한 것이다. 여기에 이후 단계에서는 Sarmatian과 한족 혈통이 더 추가되었다. 흉노의 상위 계층은 유전적 다양성이 보다 적으며, 그들의 혈통은 주로 동유라시언 Ulaanzuukh/Slab Grave culture으로부터 나왔고, 반면 하위계층의 사람들은 보다 다양하고 높은 사카 혈통을 가지는 경향이 있다. 명망있는 무덤으로 알려진 제국의 남자리더, Chanyu (선우) 같은 사람은 서쪽 국경의 상층부 여인과 유사한 혈통을 가진 것으로 나타난다. 39.3%는 Slab Grave 유전 혈통, 51.9%의 Han (한족)혈통, 그리고 나머지 8.8%의 사카 (Chandmann) 혈통을 나타낸다.
이후의 다른 동쪽으로부터의 유입은 사카의 Tasmola culture의 외지 3개 샘플과 Pazyryk culture의 하나 샘플에서 확실히 나타난다. 이들은 70-83%의 아무르 혈통을 나타냈다. 이것은 이들이 보다 동쪽으로부터 최근에 이주한 것을 말해 준다.
같은 동쪽의 혈통이 훈족의 이후그룹(Hun Berel 300년, Hun 엘리뜨 350년)과 Karakaba 유물(830년, 동카자흐스탄)에서 나타나며, 이들은 흉노족의 서쪽으로의 팽창과 관련있을 것이다. 항가리 부다페스트의 4세기 중반 엘리뜨 무덤의 훈족인은 60%의 ANA/아무르와 40%의 사카 혈통으로 이루어졌다. (아래 표 팜조)
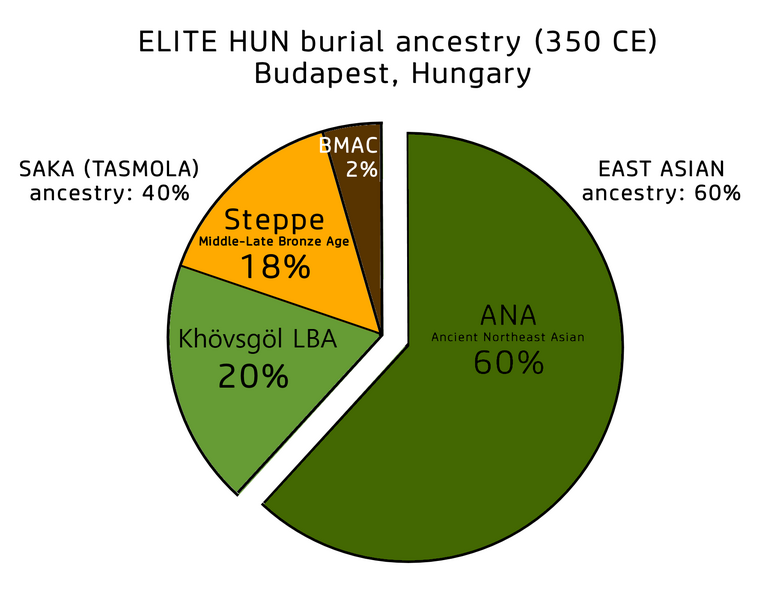
Elite Hun burial genetic ancestry (350 CE, Budapest)
A later different Eastern influx is evident in three outlier samples of the Saka Tasmola culture (Tasmola Birlik) and one of the Pazyryk culture (Pazyryk Berel), which displayed c. 70-83% additional Amur-derived ancestry, suggesting them to be recent migrants from further East. The same additional Eastern ancestry is found among the later groups of Huns (Hun Berel 300CE, Hun elite 350CE), and the Karakaba remains (830CE) and may be associated with the westwards expansion of Xiongnu tribes. A Hun individual from an elite burial of the mid-4th century CE in Budapest, Hungary, was reconstructed as 60% Ancient Northeast Asian/Amur (ANA) and 40% Saka.[40]
(번역) 이후의 다른 동쪽으로부터의 유입은 사카의 Tasmola culture의 외지 3개 샘플과 Pazyryk culture의 하나 샘플에서 확실히 나타난다. 이들은 70-83%의 추가적인 아무르 혈통을 나타냈다. 이것은 이들이 보다 동쪽으로부터 최근에 이주한 것을 말해 준다. 같은 동쪽의 혈통이 훈족의 이후 그룹(Hun Berel 300년, Hun 엘리뜨 350년)과 Karakaba 유물(830년, 동카자흐스탄)에서 나타나며, 이들은 흉노족의 서쪽으로의 팽창과 관련있을 것이다. 항가리 부다페스트의 4세기 중반 엘리뜨 무덤의 훈족인은 60%의 ANA/아무르와 40%의 사카 혈통으로 이루어졌다.
7-8세기 유럽의 아바르족은, 특히 아바르 엘리뜨에 관해서는, 주로 ANA 혈통(90%)임이 확인되었다.
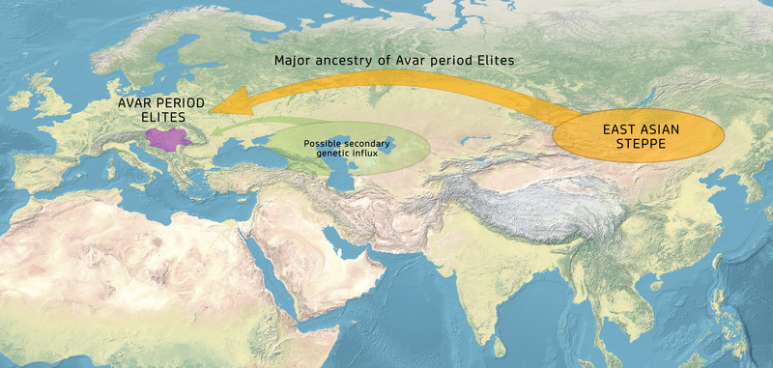
The 7-8th century Avars in Europe, particularly as regards the Avar elite, were also confirmed to have essentially Ancient Northeast Asian ancestry (c. 90%), with some additions from other sources.[41]
(번역) 7-8세기 유럽의 아바르족은, 특히 아바르 엘리뜨에 관해서는, 주로 ANA 혈통(90%)임이 확인되었다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
2. Göktürks(곡튀르크)
튀르크왕자비 Ashina(551-582)는 유전적으로 ANA(고대동북아시아인)와 밀접하게 관련되어 있는데, 97.7%의 동북아 혈통, 3천년전으로 거슬러 올라가는 2.3%의 서유라시언 혈통, 그리고 0의 지나인혈통(황하)의 혼합으로 나타난다.
이는 아시나족과 Gokturk 칸국이 동북아시아 원천임을 밑받침해 준다. 이러한 발견은 서유라시언 원천설과 여러 원천설 가정을 반박하며, 곡튀르크의 동아시아 원천을 옹호한다.
그러나 저자는 또한 튀르크제국의 사람들이 전체적으로, 특히 중앙 초원과 중세 튀르크인들은, 가변적이지만 높은 수준의 서유라시언이 혼재한다는 것을 관찰했다. 이는 제국내에서 유전적 하부구조가 있다는 것을 말해 준다. 예를 들면, 초기 중세 튀르크인들의 혈통은 유전자의 62.2%는 고대 동북아시아인(ANA), 나머지 37.8%는 서유라시언 (BMAC와 Afanasievo)로 나타나는데, 약 500년경 혼합이 이루어진 것으로 보인다. 아시나족의 튀르크족들의 지배계급은 이전의 Slab Grave 와 Ulaanzuukh culture 유해와 가까운 유전자 유사성을 나타냈다. (아래 자료 참조)
"The Turkic princess Ashina (551–582 CE), whose remains were sequenced, was found to be genetically closely associated with Ancient Northeast Asians (with 97.7% Northeast Asian ancestry, 2.3% West Eurasian ancestry dating back to around 3000 years ago, and no Chinese ("Yellow River") admixture), which according to Yang et al supports a Northeast Asian origin of the Ashina tribe and the Göktürk Khanate.[42] These findings refute "the western Eurasian origin and multiple origin hypotheses" in favor of an East Asian origin for the Göktürks.[43] However, the authors also observed that the population of the "Türkic Empire" as a whole, particularly Central Steppe and Medieval Türks, had a high but variable degree of West Eurasian admixture, suggesting genetic sub-structure within the empire:[44][45] for example, the ancestry of early medieval Turks was derived from Ancient Northeast Asians for about 62,2% of their genome, while the remaining 37,8% was derived from West Eurasians (BMAC and Afanasievo), with the admixture occurring around the year 500 CE.[46][47] The ruling clan of the Turkic peoples, the Ashina tribe, was found to display close genetic affinities with the earlier Slab Grave and Ulaanzuukh culture remains.[48]
(번역) 튀르크왕자비 Ashina(551-582)의 유전자 분자 배열 분석에 의하면, 그는 유전적으로 ANA(고대동북아시아인) (97.7%의 동북아 혈통, 3천년전으로 거슬러 올라가는 2.3%의 서유라시언 혈통, 그리고 0의 지나인혈통(황하)의 혼합)과 매우 밀접하게 연관되어 있다. 이는, Yang과 그외 사람들에 의하면, 아시나족과 Gokturk 칸국이 동북아시아 원천임을 밑받침해 준다. 이러한 발견은 서유라시언 원천설과 여러 원천설 가정을 반박하며, 곡튀르크의 동아시아 원천을 옹호한다. 그러나 저자는 또한 튀르크제국의 사람들이 전체적으로, 특히 중앙 초원과 중세 튀르크인들, 높은 정도 그러나 가변적인 수준의 서유라시언이 혼재한다는 것을 관찰했다. 이는 제국내에서 유전적 하부구조가 있다는 것을 말해 준다. 예를 들면, 초기 중세 튀르크인들의 혈통은 유전자의 62.2%는 고대 동북아시아인(ANA), 나머지 37.8%는 서유라시언 (BMAC와 Afanasievo)로 나타나는데, 약 500년경 혼합이 이루어진 것으로 보인다. 아시나족의 튀르크족들의 지배계급은 이전의 Slab Grave 와 Ulaanzuukh culture 유해와 가까운 유전자 유사성을 나타냈다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
3. ANA ancestry today
유전적으로 ANA/아무르 혈통은 현대의 동북아시아의 퉁구스어, 몽골어, Nivkh어 사용자에게서 절정에 달한다. ANA 혈통(퉁구스어 사용자 Ulchi족에 의해 대표되는)은 전반적으로 고대와 현대의 터키어, 몽골어, 퉁구스어 사용자들의 주류 혈통을 이룬다. 이것은 그들이 적어도 6천년전에 바이칼 호수지역, 동북아시아에서 서쪽으로 퍼졌음을 입증한다.
이전의 동북아시아 혈통이 시베리아로 이동하는흐름은 "신시베리아인"(우랄어를 사용하는 Nganasans에 의해 대표되는)과 관련 있으며, 이는 Yukaghir언어와 우랄어의 팽창과 관련 있으며, 약 11,000년전에 시작하는 구석기시베리아인의 부분적 이동과 관련 있는 것이다. (아래 자료 참조)
"Genetically, ANA/Amur ancestry peaks among modern Tungusic, Mongolic and Nivkh-speaking populations of Northeast Asia.[12] ANA ancestry (represented by the Tungusic-speaking Ulchi people) overall forms the main ancestry of the early and contemporary speakers of Turkic, Mongolic and Tungusic languages, which supports their spread from Northeast Asia westwards, discernable in the Lake Baikal region since at least 6kya. An earlier wave of Northern East Asian ancestry into Siberia is associated with "Neo-Siberians" (represented by Uralic-speaking Nganasans), which may be associated with the expansion of Yukaghir and Uralic languages, and the partial displacement of Paleo-Siberians, starting around 11kya.[49]
(번역) 유전적으로 ANA/아무르 혈통은 현대의 동북아시아의 퉁구스어, 몽골어, Nivkh어 사용자에게서 절정에 달한다. ANA 혈통(퉁구스어 사용자 Ulchi족에 의해 대표되는)은 전반적으로 고대와 현대의 터키어, 몽골어, 퉁구스어 사용자들의 주류 혈통을 이룬다. 이것은 그들이 적어도 6천년전에 바이칼 호수지역, 동북아시아에서 서쪽으로 퍼졌음을 입증한다. 이전의 동북아시아 혈통이 시베리아로 이동하는흐름은 "신시베리아인"(우랄어를 사용하는 Nganasans에 의해 대표되는)과 관련 있으며, 이는 Yukaghir언어와 우랄어의 팽창과 관련 있으며, 약 11000년전에 시작하는 구석기시베리아인의 부분적 이동과 관련 있는 것이다.
(source : ANA, Wikipedia, 인용출처: 필자의 네이버 블로그 (lainfos), 카테고리, 구석기 동아시아)
VI. 마무리말
이상으로 구석기, 신석기, 청동기, 철기에 걸친 동북아시아 또는 동유라시아에 대해 살펴 본 것을 마무리 하고자 한다.
전 기간에 걸친 분석 중에 특히 유난히 눈에 띄는 것은 아무르 조상이라 일컬어지는 ANA(Ancient Northeast Asians)와 Neo Siberians의 존재이다.
아무르 혈통인 ANA는 구석기시대, 26000 - 19000년전에 다른 동아시아인로부터 갈라져 나왔다는 것을 말한다. 그리고 이 아무르혈통은 그 이후 어떻게 되었는지 잘 모르나 신석기시기의 아무르조상들의 혈통이 되었다 한다. 즉 구석기시대의 고대북유라시언(ANE, 남시베리아지역)샘플(24000년전)이나 Tianyuan man 샘플(산동성지역)(4만년전)과 신석시대의 ANA샘플과는 큰 시간적 공백기간이 존재한다. 그러나 아무르지역의 AR19K(19000년전) 샘플은 신석시대의 ANA 유전인자의 토대가 된 것으로 나타난다.
특히 7000-4000년전의 수렵인그룹의 조상이며, 활동지역이 아무르지역 그리고 후에 동시베리아, 몽골, 바이칼지역에 더 퍼졌으며, 다른 고대 북쪽의 동아시아인, 예를 들어 초기 신석기 바이칼지역에 퍼진 '신시베리아인' 등에 가장 가깝게 연계되어 있다.
Ancient Northern East Asian (ANEA)은 바이칼지역부터 황하강, 그리고 현재 중공의 중앙지역인 Qinling-Huaihe Line까지 퍼져있는 고대 동북아시아인들을 대표하는 혈통이다.그들은 약 기원전 20000 - 26000년쯤에 고대 남동아시아인 (Ancient Sourhern East Asians (ASEA)로부터 갈라진 것으로 여겨진다.
고대 동남아시아인(ASEA)은, Fujian의 Qihe 동굴(12-8천년전), Guangxi(9천년전)뿐만 아니라, Taiwan 해협의 Liangdao 섬 (8천년전) 등의 인류로 대표되는 혈통계열을 말한다. ( ASEA조상은 현대의 동아시아, Mainland Southeast Asia, 섬들의 동남아시아, 그리고 오세아니아 등의 유전자 형성에 많이 기여했다.
초기 Holocene시기까지, Fujian의 고대동남아시아인(ASEA)은 유전적으로 고대동북아시아인(ANEA)과 확연히 구분된다. 이들 ASEA는 다른 동아시아의 고대 동유라시언 계열 (Tianyuan, Hoabinhian, Jomon, Guangxi/Longlin 조상들) 대비하여 확연히 구분된 계통군(clade)을 형성했다. ASEA와 ANEA계열의 분리는 적어도 19000년전에 일어났다.
중기 Holocene 시기에, ANEA 조상인자를 가진 황하의 수수 경작 농부들의 남쪽으로의 이동은 동아시아 해안의 조상인자 클라인(cline)으로 결과되어 오늘에 이르고 있다. 북쪽의 지나 한족은 대부분 적은 양의 ASEA인자와 ANEA 조상인자를 혼합하여 가지고 있다. 남쪽의 한족과 동남아시아의 비한족 그룹 (즉 Kra-Dai어 사용자와 Hmong-Mien 어 사용자)은 아직도 상당히 높은 수준의 ASEA 조상인자를 가지고 있다.
이제 신석기 시대로 넘어가 보자.
특별한 ANA유전자 그룹으로 인식되는 최초 인류는 극동러시아 태평양가에 있는 Devil's Gate Cave(기원전 5700년)에서 연유한다. ANA/아무르지역 같은 유전자그룹으로서 신석기시대 사람들은 동몽고(BC 4686-4495 BC), 중앙몽고 (BC 3781-3639 BC)에서 확인된다.
바이칼지역과 시베리아의 인접한 지역의 수렵인 그룹은, 초기 신석기의 동부 바이칼 Fofonovo culture와 서부 바이칼 Kitoi culture (5200-4200 BC)와 연관되어 있다. 뿐만 아니라, 고대 구석기시베리아인(APS)과 함께, Glazkovo culture(2500 BC)와 Cisbaikal_LNBA와 관련있는 초기 청동기시기의 바이칼 인류도 이 수렵인 그룹과 연관되어 있다.
이들 수렵인그룹은 다른 고대북동아시아인 (ANA) 그룹과 광범위하게 무리를 이루었으나, 11% (5-20%)의 작은 고대북유라시아인(ANE) 유전인자 구성에 의해 ANA그룹과 구분된다.
이러한 사람들은 때로는 '새로운 시베리아인' (Neo-Siberians)으로 묘사되며, 신석기 Devils Cave표본에 의해 대표되는 ANA/아무르 사람들과도 구분될 수 있다. 그러나 그들의 고대 동북 아시아인 (ANEA)조상을 통해 공통적인 원천을 공유하고 있다. Neo-Siberians은 신석기 아무르조상들의 팽창 이전에 이미 팽창한 것으로 여겨진다.
Devils 동굴샘플은 아무르지역의 14000년전의 샘플 (AR14K)과 유전적으로 이어지는 것을 보여 주고 있다. 이는 ANA의 특별한 유전자풀이 14000년전부터 형성되었음을 말해준다. 신석기시대의 ANA유물은 서쪽으로 1500km 떨어진 알타이산맥에서까지 발견되었다.
ANEA (Ancient Northern East Asian)는 3개의 하위그룹으로 나눌 수 있다: 즉 ANA (Ancient Northeast Asians), Neo-Siberians (신 시베리아인), 그리고 황하농민들 (Yellow River farmers) 들이다.
ANEA계열의 초기 집단은, 북으로 고대북유라시언(ANE)과 함께 고대구석기시베리아인(APS)을 형성하는데 이바지한 것으로 여겨진다. 남으로는 ASEA(고대남동아시아인)과 함께 "황하의 농민들"(Yellow River farmers)을 형성하는데 이바지 했다.
"Neo-Siberians"은, ANEA(14000년전)의 내륙확장루트와 연관 있으며, 그러나 "ANA조상"과 관련 있는 Amur-hunter-gatherers(7000-14000년전)와는 다를 수 있다.
이 Neo-Siberians은 우선적으로 신석기, 청동기시기의 바이칼지역그룹의 조상이 되었다.
이들은 초기의 범 우랄어사용자들의 팽창과 연관 있을 수 있으며, APS와 더불어, 청동기 시기 서부 바이칼 수렵인 그룹과도 연관 있을 수 있다.
고대 동북아시아인(ANA)(아무르조상)은 "Neo-Siberians"그룹이 흩어진 후 팽창했으며, 튀르크, 몽골, 퉁구스어 사용자들의 확산과 관련되어 있을 수 있다.
신석기 동남아시아인에 있어서는 쌀농사가 특징 지어진다.
쌀농사 기반의 농업은 기원전 3천년대부터, 남동아시아에서 중국 대륙 및 섬지역의 동남아시아로 퍼져 나갔다. 이러한 기술적 전파는 ASEA 조상인자를 가진 남동아시아 농업인들의 이주의 결과이다. 이러한 신석기 농부들은 두가지 길로 이동했다: 중국 본토 남동아시아로 향하는 내륙의 길, 그리고 타이완을 발생지로 하는 해상루트이다.
4천년전의 대륙 남동아시아의 최초 농부들의 고대 DNA는 ASEA계열의 혈통으로부터 비롯되는데, 지역적인 수렵인들과 상당한 혼합의 모습이다. 이 신석기 대륙의 남동아시아 조상인자는 Austroasiatic어를 사용하는 동남아시아의 그룹(특히 라오스와 타일란드의 Mlabri와 Htin 지역인들)과 동아시아, 남아시아 일부의 현대인들에게서 절정을 이룬다. 여기서, 동남아대륙의 농업의 전파는 Austroasiatic언어 사용자들의 팽창과 광범위하게 연계되어 있다.
5천-4천년전부터 시작한 초기 Austronesians의 해상을 통한 팽창은 타이완부터 필리핀, 인도네시아 군도, 오세아니아로 ASEA 조상인자가 퍼졌다. 이후 Austronesian 언어 사용자들의 이동은 ASEA조상인자를 멀리 마다가스카르와 동부 폴리네시아까지 이동시켰다.
청동기에 들어서는 동유라시아 바이칼호수와 몽고지방의 활동, 즉 사카의 활동이 두드러진다.
Ulaanzuukh(1450-1150 BC)와 Slab Grave (1100-300 BC) 문화의 사람들은 ANA(아무르 조상인자)의 직접 자손들로 정형화할 수 있다. 그들은 이전의 신석기 및 초기 청동기시기의 바이칼 수렵인들을 광범위하게 대체했다. 그들 사이의 유전적 교류가 있었는데, 특히, 원래 Amur-N과 같은 혈통을 가진 신석기 동부 몽골인들과 지역의 바이칼 수렵인들 사이의 유전적 교류가 있었다 한다.
신석기 및 초기 청동기시기 바이칼 수렵인 후계 그룹은 서부초원유목민(Western Stepple Herders)/Sintashta 문화 같은 이들과 다양한 정도의 혼합이 후기 청동기시기동안에 알타이지역에서 나타나기 시작했다. 이러한 그룹은 동부초원으로부터 신석기 및 초기 청동기 바이칼지역인들로부터 형성되었다. 그리고 서부초원유목민(WSH) 이주그룹과의 연이은 혼합이 이루어졌다. 이것은 북몽골의 Khovsgol LBA (Late Bronze Age)유목민과 알타이지역의 Altai MLBA (Middle Late Bronze Age)수렵인그룹을 포함한다.
Khovsgol (바이칼호수 중간 아래) 후기청동기 유목민들은 대부분의 초기 청동기 바이칼 수렵인그룹(Baikal EBA or Shamanka EBA, 93-96%)과 약간의 서부초원유목민(WSH) (Sintashta, 4-7%)들의 혼합의 자손들이다. 유전적 분석은, 이들이 서부초원유목민들(WSH)로부터 낙농유목이 채택되었고, 그들은 원래 지역의 동북아시아 출신임이 밝혀졌는데, 이는 문명적 전파를 의미한다
더 서쪽의 알타이 중기후기청동기시기 유전적 그룹은, 동부 스키타인(사카) (바이칼 /Shamanka EBA그룹의 40-55% 조상인자를 가진 그룹)과 연관되어 있으며, 또한 나머지부분은 초기 인도이란인과 관련있는 Sintashta같은 그룹과 혼합 (45-60%)되어 있다.
타림 미이라(기원전 2000년)는 높은 수준의 ANE(고대 북유라시언) 조상인자(72%)와 보다 적은 동아시아인구 (특히 Baikal_EBA 28%)의 혼합을 보여 주었다.
철기 동유라시아는 사카 흉노제국, 훈제국, 아바르제국, 곡튀르크 등으로 특징지어진다.
바이칼 초기 청동기 인류는 혼종의 유라시언 Scytho-Siberians cultures의 형성에 지대한 공헌(40-55%)을 하였다. 이들 Scytho-Siberians cultures는 BC 1000년경부터 중앙아시아의 Tasmola문화 (Central Saka) 뿐만 아니라, Arzhan, Pazyryk culture (동부 사카)들을 말한다.
혼종의 사카문화는 흉노제국 (기원전 3세기-기원후 1세기)의 형성에 중요한 역할을 했다. 이 흉노제국은 특별한 사카 혈통(특히 Chandman/Uyuk같은 것들)과 신석기 아무르 혈통 (Ulaanzuukh 와 Slab Grave 혈통들)을 합한 것이다. 여기서 흉노제국은 중공사학자들이 주장하는 중공 북방의 민족을 말한다. 이후에는 Sarmatian(동유럽)과 한족 혈통이 더 추가되었다.
흉노(=중공 북방유목민)의 상위 계층은 유전적 동질성이 보다 많으며, 그들의 혈통은 주로 동유라시언 (Ulaanzuukh/Slab Grave culture)으로부터 나왔다. 반면 하위계층의 사람들은 보다 다양하고 높은 바율의 사카 혈통을 가지는 경향이 있다.
제국의 남자리더, 'Chanyu (선우)' 같은 사람은 서쪽 국경의 고위계층 여인과 유사한 혈통을 가진 것으로 나타난다. (중공북방의 흉노 유목민은) 39.3%는 Slab Grave 유전 혈통, 51.9%의 Han (중공지역 한족)혈통, 그리고 나머지 8.8%의 사카 (Chandmann) 혈통을 나타낸다.
이후의 다른 동쪽으로부터의 유입은 사카의 Tasmola culture의 외지 3개 샘플과 Pazyryk culture의 하나 샘플에서 확실히 나타난다. 이들은 70-83%의 아무르 혈통을 나타냈다. 이것은 이들이 보다 동쪽으로부터 최근에 이주한 것을 말해 준다.
같은 동쪽의 혈통이 훈족의 이후그룹(Hun Berel 300년, Hun 엘리뜨 350년)과 Karakaba 유물(830년, 동카자흐스탄)에서 나타나며, 이들은 흉노족의 서쪽으로의 팽창과 관련있을 것이다. 항가리 부다페스트의 4세기 중반 엘리뜨 무덤의 훈족인은 60%의 ANA/아무르와 40%의 사카 혈통으로 이루어졌다.
7-8세기 유럽의 아바르족은, 특히 아바르 엘리뜨에 관해서는, 주로 ANA 혈통(90%)임이 확인되었다.
튀르크왕자비 Ashina(551-582)는 유전적으로 ANA(고대동북아시아인)와 밀접하게 관련되어 있는데, 97.7%의 동북아 혈통, 3천년전으로 거슬러 올라가는 2.3%의 서유라시언 혈통, 그리고 0의 지나인혈통(황하)의 혼합으로 나타난다.
이는 아시나족과 Gokturk 칸국이 동북아시아 원천임을 밑받침해 준다. 이러한 발견은 서유라시언 원천설과 여러 원천설 가정을 반박하며, 곡튀르크의 동아시아 원천을 옹호한다.
그러나 튀르크제국의 사람들이 전체적으로, 특히 중앙 초원과 중세 튀르크인들은, 높은 수준의 서유라시언이 혼재한다. 이는 제국내에서 유전적 하부구조가 있다는 것을 말해 준다. 예를 들면, 초기 중세 튀르크인들의 혈통은 유전자의 62.2%는 고대 동북아시아인(ANA), 나머지 37.8%는 서유라시언 (BMAC와 Afanasievo)로 나타나는데, 약 500년경 혼합이 이루어진 것으로 보인다. 아시나족의 튀르크족들의 지배계급은 이전의 Slab Grave 와 Ulaanzuukh culture 유해와 가까운 유전자 유사성을 나타냈다.
유전적으로 ANA/아무르 혈통은 현대의 동북아시아의 퉁구스어, 몽골어, Nivkh어 사용자에게서 절정에 달한다. ANA 혈통(퉁구스어 사용자 Ulchi족에 의해 대표되는)은 전반적으로 고대와 현대의 터키어, 몽골어, 퉁구스어 사용자들의 주류 혈통을 이룬다. 이것은 그들이 적어도 6천년전에 바이칼 호수지역, 동북아시아에서 서쪽으로 퍼졌음을 입증한다.
이전의 동북아시아 혈통이 시베리아로 이동하는흐름은 "신시베리아인"(우랄어를 사용)과 관련 있으며, 이는 Yukaghir언어와 우랄어의 팽창과 관련 있으며, 약 11,000년전에 시작하는 구석기시베리아인의 부분적 이동과 관련 있는 것이다.
아무르 혈통인 ANA그룹이 멀리 서쪽으로 간, 훈족이나 아바르족, 그리고 곡튀르크와 유전적으로 관련 있다는 분석은 매우 시사하는 바가 크다. 동북아시아에서 대서양까지 진출했다는 결론이 되기 때문이다. 필자의 그동안의 역사 공부가 허사가 아니라는 것을 보여 준다. 그러나, 유럽으로의 이동이 ANE의 이동역사 같이 14000년전 - 9000년전 기간이나 또는 철기시대에 들어 확인되는 것으로 보인다. 따라서 12환국의 위치를 동북아시아에 한정하는 것은 합리적이지 않은 것으로 판단된다. 동북아시아에서도 홍산문화, 황하의 앙소문화가 연결된 모습을 필자는 지난 글에서 살펴 보았다. 따라서 신석기 시기의 바이칼호수, 몽고지역의 일부 사카 활동은 12환국의 위치를 합리화 하기 충분하지 않다.
다음에는 유럽으로 간 스키타이(서 사카)와 동부사카의 역사를 보면서 12환국의 위치를 가늠해 보고자 한다.
'한국사 > 기타' 카테고리의 다른 글
| 파내류산은 북유럽에 있었다 (1) | 2023.12.07 |
|---|---|
| 흉노의 고향(=약수)은 어디인가? (0) | 2023.10.30 |
| 돌궐, 회흘은 북해 위에 있었다 (0) | 2023.09.06 |
| 네르친스크 조약이 언급하는 아무르강은 아무다리야강이 아닐까? (2) (2) | 2023.06.05 |
| 네르친스크조약이 언급하는 아무르강은 아무다리야강이 아닐까? (I) (0) | 2023.05.26 |